 Variations
in peat bog spider communities related to environmental heterogeneityVariations
in peat bog spider communities related to environmental heterogeneity
Variations
in peat bog spider communities related to environmental heterogeneityVariations
in peat bog spider communities related to environmental heterogeneityThe composition and structure of araneid/opilionid communities depends upon environmental conditions in complex ways. Firstly, habitat properties determine the distribution and thus the probability of co-occurrence of species to form a community. Secondly, community structure is the expression of component species' interactions, mostly competitive, which are influenced by environmental patchiness and the distribution of species over microhabitats. In this paper we are concerned with data illustrating the influence of environmental heterogeneity on the ground-layer spider (and harvest-spider) communities of peat bogs.
Vegetation physiognomy is a complex indicator of habitat conditions. The physical structure of vegetation is important in itself, but it also depends upon and influences microhabitat conditions. Plant tussocks or the dwarf shrubs which are found on peat bogs provide shelter with relatively constant microclimates affected by the height, density and orientation of the vegetation. Vegetation physiognomy on the peat bogs is also related to many other aspects, such as general climatic conditions, degree of exposure, peat drainage, and other effects of management, e.g. periodic moor-burning. Phytosociological characteristics could also be taken into account and will be considered elsewhere.
Single sample stations were placed on Blood Moss, Dalmellington Moss and Longriggend Moss, but at the partially wooded bogs samples were taken in both open and wooded areas. Carnwath Moss and Wood, Braehead Moss and Wood, North Bellstane Plantation (NBP) Moor and Wood, Bloak Moss and Wood. Birch woodland is present at Braehead and NBP Woods, the latter having a very wet floor and relatively lush vegetation. In contrast, Bloak Wood is an anthropogenic Scots pine plantation with ground flora formed by degeneration of the peat-bog community drying out. At Carnwath the Wood site is in sparse birch and pine on a drier area of the bog with degeneration and consequent patchiness of the heather.
Ten pitfall traps with formalin/detergent trapping fluid were set in a 1 m2 quadrat at each sampling station. These were emptied at varying intervals and data combined over the whole year; temporal variation is not considered here. Environmental variability of each site was assessed by examining the vegetation at 1 m intervals over a 5 x 5 m grid centred on the pitfall station.
Contributors |
Principal Components |
|||
Features |
Height Range (cm) |
I |
II |
III |
| Vertical | 0 - 10 | 0 | -0.457 | 0.558 |
| 10 - 20 | 0.375 | -0.316 | 0.196 | |
| 20 - 30 | 0.449 | 0.179 | 0 | |
| 30 - 40 | 0.263 | 0.356 | 0.184 | |
| 40 - 50 | 0.113 | 0.256 | 0.235 | |
| 50 - 60 | 0 | 0.122 | 0 | |
| Horizontal | 0 - 10 | 0.188 | 0 | -0.551 |
| 10 - 20 | 0.246 | 0 | -0.406 | |
| 20 - 30 | 0.184 | 0.103 | 0 | |
| 30 - 40 | 0 | 0.136 | 0 | |
| Density | 0 - 10 | 0.238 | -0.443 | 0 |
| 10 - 20 | 0.409 | -0.234 | -0.122 | |
| 20 - 30 | 0.383 | 0.152 | 0 | |
| 30 - 40 | 0.209 | 0.258 | 0 | |
| 40 - 50 | 0 | 0.207 | 0.161 | |
| % of total variance:- | 43.8 | 21.5 | 9.4 | |
| Interpretation:- | abundance | shrubbiness | orientation | |
| all non-zero values
are positive |
contrast negative
or zero values near ground with positive values higher up |
contrast positive
values for vertical features with negative values for horizontal |
||
The PCA plot in Fig. 1 indicates the relative positions of the sites,
in terms of vegetation structure, and their different amounts of variation.
The latter may be quantified by the trace (T) of the covariance matrix,
i.e. the summed variances of all variables. The volume (V) enclosed by
an ellipsoid described by taking one standard deviation to either side
of a site's centroid of PCA scores may be used to express the relative
spread of the site in a hyperspace defined by the PCA axes. Values of T
and V are given in Table 2; they represent measures of environmental heterogeneity.
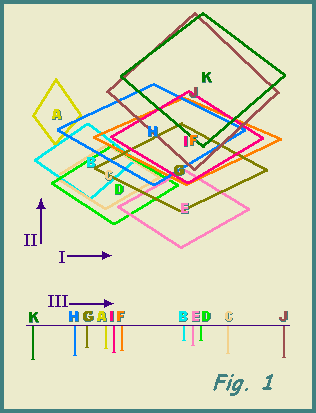
Fig. 1. Vegetation physiognomy PCA.
Above: plot of axes I and II.
Letters indicate the centroids of scores for 25 samples at each site
A - K and are surrounded by lines drawn to apices at one standard deviation
either side of mean on each axis.
Arrows indicate increasing vegetation density a1ong axis I and increasing
shrubbiness along axis II,
Below: sites along axis III.
Arranged from predominantly horizontal to mainly vertical orientation
of structures. Lengths of lines drawn proportional to standard deviation
of scores at each site.
| Site | T | V | N | S | E(S) | B |
| A Braehead Wood | 199 | 915 | 939 | 59 | 28.8 | 16.48 |
| B Longriggend Moss | 233 | 1401 | 1247 | 60 | 26.0 | 10.12 |
| C Dalmellington Moss | 323 | 2041 | 686 | 45 | 23.8 | 7.80 |
| D Blood Moss | 339 | 1833 | 886 | 35 | 16.1 | 6.46 |
| E Carnwath Moss | 356 | 2046 | 1036 | 61 | 28.6 | 15.17 |
| F Braehead Moss | 516 | 4355 | 862 | 54 | 29.6 | 11.95 |
| G NBP Wood | 641 | 3972 | 989 | 72 | 37.0 | 20.95 |
| H Bloak Wood | 699 | 6721 | 790 | 65 | 31.7 | 13.58 |
| I Bloak Moss | 733 | 5764 | 401 | 49 | 30.3 | 12.37 |
| J NBP Moor | 949 | 10189 | 1147 | 63 | 30.2 | 14.92 |
| K Carnwath Wood | 996 | 10085 | 867 | 66 | 35.4 | 26.00 |
A PCA plot of the spider data is shown in Fig. 2, on which deduced gradients
are indicated. 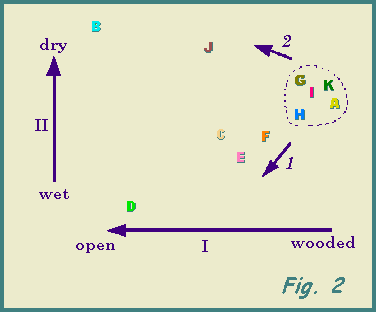
The first three axes accounted for 32.6%, 20.3% and 11.9% respectively
of the total variance.
Discussion is here confined to the more abundant species and the distribution
of individual species will be considered fully elsewhere. Relative contributions
made by the various species indicates groupings:- (a) a woodland
or litter-dwelling group includes opilionids plus Diplocephalus picinus
(Blackwall 1841), Lepthyphantes zimmermanni Bertkau 1890, Pardosa
nigriceps (Thorell 1856), Pirata uliginosus (Thorell 1856),
Gonatium
rubens (Blackwall 1833) and Lepthyphantes tenebricola (Wider
1834); (b) Alopecosa pulverulenta (Clerck 1757) and
Pachygnatha
degeeri are associated with dry, open sites, while (c) Pardosa
pullata (Clerck 1757),
Pirata piraticus (Clerck 1757) and Centromerita
concinna Thorell 1875 tend to wetter, open sites;
(d) Lepthyphantes
mengei Kulczynski 1887, Cnephalocotes obscurus (Blackwall 1834),
Trochosa
terricola Thorell 1856 and
Antistea elegans are intermediate
between group (a) and groups (b) and (c).
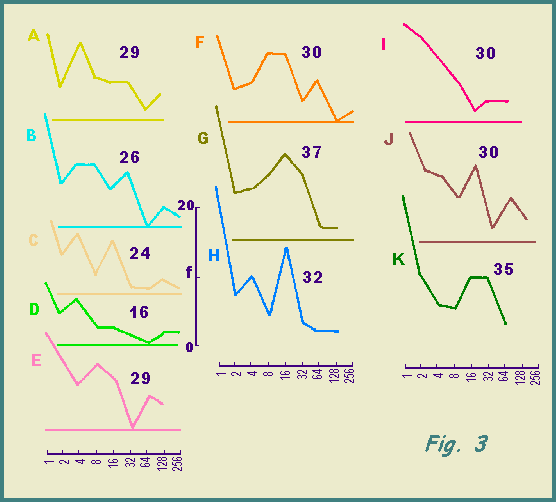
The value of E(S) depends upon the frequency of species represented by different numbers of individuals and species frequency curves are drawn in Fig. 3. Variations are apparent from one site to another.
| T | V | N | S | E(S) | B | |
| T | 0.964 | -0.218 | 0.509 | 0.746 | 0.427 | |
| V | 0.000 | -0.277 | 0.491 | 0.691 | 0.355 | |
| N | - | - | 0.382 | -0.136 | 0.236 | |
| S | - | - | - | 0.782 | 0.809 | |
| E(S) | 0.008 | 0.018 | - | 0.005 | 0.746 | |
| B | - | - | - | 0.003 | 0.008 |
There is good agreement between the community parameters (highlighted yellow) and also between V and T (highlighted green); N does not correlate with anything. Without the two birch-wood sites, there is a correlation of 0.683 between S and V (p = 0.041), i.e. between species richness and environmental heterogeneity. The pink cells in Table 3 highlight the significant correlation between the "expected number of species" and environmental heterogeneity.
Relatively few species were recorded at site D, at a higher altitude and more exposed than other sites, but relatively many at the two birch-wood sites, A and G, influenced by the occasional migration of canopy and field-layer species to the ground.
Because of the mobility of spiders, it is difficult to distinguish 'between-habitat' from 'within-habitat' diversity. In other words, are we measuring point-, or α-diversity, sensu Whittaker (9), or is it so affected by movement of species from adjacent (micro-) habitats that it represents β-diversity? A relationship between species diversity and spatial variation has been hypothesised (7) for spider communities and demonstrated for many other groups of animals. Increased environmental heterogeneity permits the coexistence of more species (6,8) and may influence the competitive interactions in the community (5).
Pianka (4, p.292) neatly lists ten mechanisms which may influence species diversity. Given that these interact with each other, it is clear that the data reported here agree with the spatial heterogeneity hypothesis, but probably are not entirely explained by it alone.
(2) CURTIS, D.J. (1980): A simple method for obtaining quantitative information on vegetation physiognomy. Scottish Field Studies, 1980, 5 - 11.
(3) HECK Jr., K.L , VAN BELLE, C. & SIMBERLOFF, D. (1975): Explicit calculation of the rarefaction diversity measurement and the determination of sufficient sample size. Ecology 56, 1459-1461.
(4) PIANKA, E.R. (1978): Evolutionary Ecology. New York (Harper & Row).
(5) POST, W.M. & RIECHERT, S.E. (1977): Initial investigations into the structure of spider communities. I. Competitive effects. J. Anim. Ecol. 46, 729-749.
(6) TURNER, M. & POLlS, C.A. (1979): Patterns of coexistence in a guild of raptorial spiders. J. Anim. Ecol. 48, 509-520.
(7) UETZ, G.W. (1975): Temporal and spatial variation in species diversity of wandering spiders (Araneae) in deciduous forest litter. Environ, Entomol. 4, 719-724.
(8) UETZ, G.W. (1977): Coexistence in a guild of wandering spiders. J. Anim, Ecol. 46, 531-541.
(9) WHITTAKER, R.H. (1969): Evolution of diversity in plant communities. Brookhaven Symp. Biol. 27, 178-196.
Dr. E. Bignal, c/o Joint Nature Conservation Committee, Northminster House, Peterborough, UK.
.
| Return to..? | |
 Ariadne Home Page
Ariadne Home Page |
|