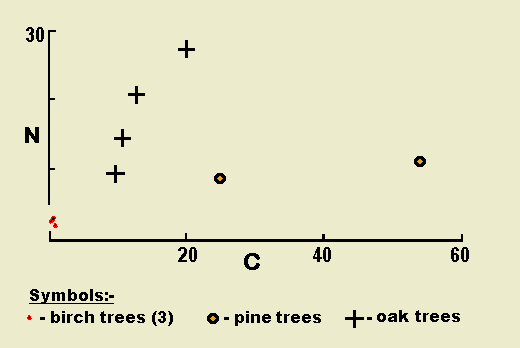
The numbers of individuals of spiders trapped on tree trunks appear to show a correlation with the degree of cracking af the bark for the same species of tree, but vary from one kind of tree to another.
Two relatively simple measures of diversity are illustrated which yield a quantitative estimation of the dispersion of a species between different habitats.
A simple measure of overlap is illustrated which gives a quantitative impression of the extent to which two species share their habitat ranges.
The work described here was carried out in woodland on the island of Inchcailloch in the Loch Lomond National Nature Reserve. This island is located about 500m off Balmaha Bay and is approximately 1400m long with a maximum width of 600m. Because of the absence of grazing stock since the end of the last century, the island bears some of the best natural deciduous woodland remaining in Scotland today, consisting predominantly of oak.
The traps used to sample the tree trunk populations were of the corrugated paper type which Duffey (1969) used to investigate seasonal movements of clubionids. Nine trees were sampled - 3 mature and 1 young oak (Quercus sp.), 3 birch (Betula pubescens), 2 Scots pine (Pinus sylvestris) - located at five sites on the island. On each tree a strip of brown corrugated paper approximately 22cm wide was wrapped twice around the trunk about 1.5m above the ground. Each trap was held in place by string and two nails upon which it rested to prevent it slipping down the trunk. In addition, 10cm wide grease bands coated with adhesive resin were wrapped around the outside of the traps. The corrugations in the paper provided refuges for many small invertebrate animals and presumably function as extra "cracks and crevices in the bark". Each tunnel formed by the corrugations measured about 3 x 3 mm across; this imposes a maximum size limit on spiders taking shelter in the paper.
At weekly or fortnightly intervals, the corrugated paper traps and grease bands were removed and replaced by fresh ones. Each old trap was returned to the laboratory along with any disturbed specimens. Removal of invertebrates followed anaesthetization with chloroform and the careful stripping of the backing paper to expose the tunnels. Those arachnids evident on the grease bands were extracted and cleaned with acetone. The spiders were identified using the keys of Locket and Millidge (1951, 1953) and nomenclature follows theirs. The phalangids and pseudoscorpions were determined with the aid of Savory (1948) and Evans and Browning (1954) respectively.
The captures from the traps are listed in Table 1. A total of 22 species of spiders were taken, as well as some specimens rendered unidentifiable by immaturity or damage. Two species of phalangids and one pseudoscorpion species were also taken. Nineteen aranaeid species were found in the corrugated paper; of these species, six were also taken on the grease bands. The grease band captures included nine spider species. There was a marked difference in the sex ratios of the adult spiders caught in the corrugated paper traps (sex ratio O.6 males : 1 female) and those from the grease bands (5.5 males : 1 female). Possibly this reflects a greater tendency on the part of the males to move about more on the exposed surfaces of the bark and for the females to remain sheltered in the cracks.
| Table 1: List of captures from corrugated paper traps and grease bands. | |||||
|---|---|---|---|---|---|
| Numbers of arachnids captured in corrugated paper traps and on grease bands on trees at the five different sites. Numbers in brackets refer to individuals displaced from traps on removal, counted but not caught. (Site descriptions at foot of table.) | |||||
| Species | Corrugated paper | Grease bands | |||
| Nos. | Sites | Nos. | Sites | ||
| Ciniflo fenestralis (Stroem) | 3♂ 19♀ 50imm(4) | 1,2,3 | 1imm | 2 | |
| Harpactea hombergi (Scopoli) | 2♂ 3♀ | 1,2 | |||
| Segestna senoculata (Linn.) | 1♀ | 2 | |||
| Clubiona compta C.L.Koch | l♂ 2imm | 2 | 1♀ | 2 | |
| Anyphaena accentuata (Walckenaer) | 3♂ 4♀ 2imm(1) | 1,3,4,5 | |||
| Xysticus (lanio?) C.L.Koch | 1imm | 1 | |||
| thomisid (damaged) | 1♀ | 5 | |||
| Cryphoeca silvicola (C.L.Koch) | 3♂ 46♀ 18imm | 1,2,4 | 12♂ 5imm | 1,4 | |
| Robertus lividus (Blackwall) | 1imm | 4 | |||
| Tetragnatha montana Simon | 2♀ | 1,4 | 1imm | 2 | |
| Tetragnatha (obtusa?) C.L.Koch | 2imm | 2 | |||
| immature argiopid | 1 | 1 | |||
| Araneus umbraticus Clerck | 2♀ | 1,2 | |||
| Ceratinella brevipes (Westring) | 1♂ | 4 | |||
| Gonatium rubellum (Blackwall) | 1♀ | 4 | |||
| Lophocarenum nemorale (Blackwall) | 1♀ | 1 | |||
| Thyreosthenius parasiticus (Westring) | 1♂ 2♀ | 3,4 | |||
| Monocephalus fuscipes (Blackwall) | 1♂ 2♀ | 4 | |||
| Savignia frontata (Blackwall) | 2♂ | 2 | |||
| Diplocephalus cristatus (Blackwall) | 6♂ 1♀ | 4 | |||
| Caledonia evansi O.P.-Cambridge | 2♀ | 4 | |||
| Drapetisca socialis (Sundevall) | 1♂ 3♀ | 1,2 | 1♀ | 2 | |
| Lepthyphantes alacris (Blackwall) | 1♀ | 2 | 1♀ | 1 | |
| Linyphia peltata Wider | 2♂ 1♀ 5imm | 1,4 | |||
| immature linyphiids | 3 | 1 | 6 | 1,2,3 | |
| unidentifiable (damaged or imm.) | 4 | 1,2,4 | 21 | 1,2,3,4 | |
| Megabunus diadema (Fabricius) | 9 | 1,2,5 | |||
| Oligolophus hanseni (Kraepelin) | 1 | 2 | |||
| Chthonius (tetrachelatus?) (Preyssler) | 3 | 3 | |||
| Site 1 - in open woodland; some bracken and bramble; altitude 40m; soil: brown podzol on serpentine rock. Site 2 - on North Hill amongst Scots pine; some bracken, heather and blaeberry; altitude 61m; soil: peaty ranker on sandstone. Site 3 - exposed on burnt heather and tufted grass on Tom Na Nigheann summit; altitude 85m; soil: as site 2. Site 4 - in South Central Valley in deciduous woodland with oak, hazel, bramble, bracken; altitude 27m; soil: brown podzol on sandstone and conglomerates. Site 5 - near north coast of island, 8m from lochside in open oak woodland with bramble; altitude 9m; soil: gleys on mixed colluvium and drift. | |||||
Of the 19 species of spiders taken in the corrugated paper traps, two were very abundant. The relative proportions of six of the species are given in Table 2. Ciniflo fenestralis (29.56% of total) and Cryphoeca silvicola (39.86%) together constitute nearly 70% of the total catch. The next most plentiful species was Anyphaena accentuata which formed about 8% of the catch; the remaining species were taken only relatively rarely, their proportional representations being only of the order of 0.5-1.5%. These percentages are based on flgures adjusted to a unit area of trap per week; a procedure made necessary the different girths of the trees sampled.
| Tab1e 2: Relative proportions and diversity indices for some spiders taken in corrugated paper traps. | ||||
|---|---|---|---|---|
| Species No. | Species | % of total | B | B |
| 1 | Ciniflo fenestralis | 29.56 | 4.26 | 0.64 |
| 2 | Segestria senoculata | O.44 | 1 .00 | 0 |
| 3 | Anyphaena accentuata | 8.21 | 6.36 | 0.81 |
| 4 | Cryphoeca silvicola | 39.86 | 3.01 | 0.55 |
| S | Drapetisca socialis | 1.68 | 2.03 | 0.30 |
| 6 | Lepthypharntes alacris | 0.22 | 1.00 | 0 |
| Percentage of catch, Levins index of diversity (Bi) and Shannon index (B Based on figures adjusted to unit area of trap per week. |
||||
Again using figures adjusted to unit area of trap per week, the catch from the different trees varies widely. The crack areas of the sampled trees were estimated by measuring the area of cracks in a quadrat 20cm x 20cm square at the position of the corrugated paper trap. There appears to be some correlation between the amount of cracking of the bark of oaks and the number of spiders. The mature oak trees, with well cracked bark, provided most of the spider captures. The pine trees, although with a much more broken surface texture than the oaks, yielded fewer spiders. The birch trees with their relatively smooth surface gave very few spiders in the traps. These results are illustrated in Figure 1, in which the oaks show an increase in the number of spiders caught with increasing cracking of the bark.
|
The small number of trees sampled during this brief project do not make the results amenable to statistical treatment, but the results nevertheless indicate a possible interesting relationship worth further study.
Niche breadth, or versatility, is defined (Colwell & Futuyma, 1971) in spatial model terms as the "distance through" a niche along some particular line in niche space. Essentially, niche breadth may be regarded as the inverse of ecological specialisation. A spider, for example with very specialised eating habits will have a narrower niche breadth than a more general eater. Much has been written on diversity as an ecological concept, for example Pielou (1966, 1969, 1972), Levins (1968), McIntosh (1967), Hurlbert (1971), Margalef (1958), and Simpson (1949).
As examples of such indices, two are applied to the data obtained from the corrugated paper traps. The figures from traps on the different trees can be used to determine the dispersion of each species amongst these different habitats. A table, the resource matrix in the terminology of Colwell & Futuyma (1971) is drawn up with species as rows and traps ("resource states") as columns. The number of individuals per unit area (500cm2 ) of trap per week was calculated and set down in the resource matrix. For species i associated with resource state j, this number is designated Nij. The total number of individuals of species i ( the row total = Σj Nij ) is called Yi . The dispersion of a species in the resource matrix can be estimated by measuring the uniformity of the distribution of individuals of that species among the resource states of the matrix (i.e. among the traps).
Two simple measures of uniformity are Levin's (1968) formula:
Bi = Yi2 / Σj Nij2
and the Shannon-Weaver (1963) formula for information content:
Bi' = - Σj pij log pij = Σj (Nij/Yi) log(Yi/Nij);
pij = Nij/Yi is the proportion of individuals of species i associated with resource state j.
Σj means sum over all values of j.
B
Far each of the six selected species, the two different indices are listed in Table 2. According to these results the greatest dispersion is shown by Anyphaena accentuata (B = 6.36 B' = 0.81). Members af this species were found on seven of the nine trees sampled, on all three types of tree (oak, pine and birch), and at all five sites. Ciniflo fenestralis (B = 4.26; B' = 0.64) and Cryphoeca silvicola (B = 3.01; B' = 0.55) showed the next highest dispersions, even though these species were far more numerous than A. accentuata. There is good agreement between the order of values for both indices, although their actual ranges differ.
Comparison of the distribution of species can be a rather subjective process, but can be rendered objective by the utilisation of a quantitative measure of overlap (or similarity). Some such measures are discussed by Horn (1966) and they can be quite difficult to calculate without the aid of a computer, e.g. that of Morisita (1959) which is based on Simpson's (1949) index of diversity. One discussed by Colwell and Futuyma (1971), and elaborated by them, is relatively simple to calculate and is applied to the corrugated paper trap data here.
The index of overlap used here to compare species is given as:
Cih =
1 - ½∑j| pij - phj |
where pij and phj refer
to the proportions of species i and h respectively in habitat j. | pij - phj | is the absolute difference between these, irrespective of sign. The values of C, corrected to two decimal places and multiplied by 100 to give percentage overlap, are given in Table 3.
| Table 3: Indices of overlap far some spiders. | ||||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Spiders taken in corrugated paper traps. Index of overlap expressed as percentage. | ||||||||||||||||||||||||||||
| ||||||||||||||||||||||||||||
| Species numbers as in table 2. |
Values range from zero for no overlap to 100 for complete overlap in distribution of the species. Such a measure is more useful in the case of species pairs where C lies away from the extreme values. In the present case, for species which were taken on only one and the same tree, they obviously completely overlap and the calculated value of C is in fact 100, e.g. species numbers 2 and 6. Similarly those species taken rarely but from different trees have an overlap of zero, e.g. 2 and 3, 2 and 4. The three most abundant species - 1 (C. fenestralis), 3 (A. accentuata) and 4 (C. silvicola) - show rather lower overlap than one might have expected on a subjective estimation:-
C1,3 = 38; C1,4 = 36; C3,4 = 48.
Such a measure of overlap is thus a useful tool for application to any set of results for distribution of species between different habitats. It can also be utilised in an analogous mannner to compare the species compositions of faunas at different sites.
An alternative to the calculation of overlap of the species' distributions is the computation of a distance measure such as
Dih = √ Σj (nij - nhj)2 , where nij and nhj are the numbers of individuals of species i and h at site j (McIntosh, 1967). This is related to the diversity index proposed by McIntosh (1967) which is given by √ Σi ni2, where ni is the number of individuals of each species i at the site. These latter indices are computed on the basis of the actual numbers of individuals taken, rather than the relative proportions of the species and are thus sensitive to the abundances of the species at the differerit sites. The relationships between various indices have been studied by DeBenedictis (1973). The distribution of values of diversity indices with communities of different numbers of individuals and of species is a complex problem and the interested reader is referred to the work of Bowman et al. (1971) for a detailed mathematical treatment.
The authors are grateful to Dr J.C. Smyth and Mr. J.D. Hamilton of the Department of Biology, Paisley College of Technology, for help and useful discussions. We also thank members of the Nature Conservancy for permission to work on the island and other assistance, especially Mr. E. Idle, Mr C. Placido and Mr J. Mitchell.
BOWMAN, K.G., HUTCHESON, K. ODUM, E.P. & SHENTON, L.R. 1971: Comments on the distribution of indices of diversity. Statistical Ecology vol. 3. Many species populations, ecosystems and systems analysis. (ed. Patil, G.P., Pielou, E.C. & Waters, W.E.)
COLWELL, R.K. & FUTUYMA, D.J. 1971: On the measurement of niche breadth and overlap. Ecology 52: 567-576
DeBENEDICTIS, P.A. 1973: On the correlation between certain diversity indices. Am. Nat. 107: 295-302
DUFFEY, E. 1969: The seasonal movements of Clubiona brevipes Blackwall and Clubiona compta C.L.Koch on oak trees in Monks Wood, Huntingdonshire. Bull. Br. Arach. Soc. 1: 29-32
EVANS, G.O. & BROWNING, E. 1954: Pseudoscorpiones. Synopses of the British Fauna No. 10. London, Linnean Society of London.
HORN, H. 1966: Measurement of "overlap" in comparative ecological studies. Am. Nat. 100: 419-424
HURLBERT, S.H. 1971: The nonconcept of species diversity: a critique and alternative parameters. Ecology 52: 577-586
LEVINS, R. 1968: Evolution in changing environments. Princeton, Princeton University Press
LOCKET, G.H. & MILLIDGE, A.F. 1951: British Spiders. Vol. 1. London, Ray Society
LOCKET, G.H. & MILLIDGE, A.F. 1953: British Spiders. Vol. 2. London, Ray Society
MARGALEF, D.R. 1958: Information theory in ecology. Gen. Syst. 3: 36-71
McINTOSH, R.P. 1967: An index of diversity and the relation of certain concepts to diversity. Ecology 48: 392-404
MORISITA, M. 1959: Measuring of interspecific association and similarity between communities. Mem. Fac. Sci., Kyushu Univ. Ser. E. (Biology) 3 65-80
PIELOU, E.C. 1966: The measurement of diversity of different types of biological collections. J. theoret. Biol. 13: 131-144
PIELOU, E.C. 1972: Niche width and niche overlap: a method for measuring them. Ecology 53: 687-692
SAVORY, T.H. 1948: Opiliones. Synopses of the British Fauna No. 1. London, Linnean Society of London
SHANNON, C.E. & WEAVER, W. 1963: The mathematical theory of communication. Urbana, University of Illinois Press
SIMPSON, E.H. 1949: Measurement of diversity. Nature, Lond. 163: 688
 Ariadne Home Page Ariadne Home Page | |