Ground fauna was sampled at four sites in woodland on the island of Inchcailloch. About 15000 adults of 11 species of harvestmen and 101 species of spiders were recorded.
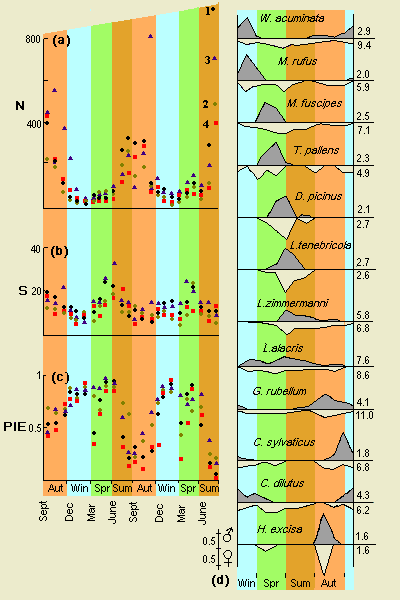
Relative abundance at four sites was in order: 1,3,4,2. Species richness was in order: 3, 1, 2,4. Species diversity (PIE) was in order: 2, 3, 4, 1.
Number of individuals (N) caught were maximal in late summer and
early autumn, when PIE values were lowest. PIE and N were negatively
correlated, but species richness independent of N.
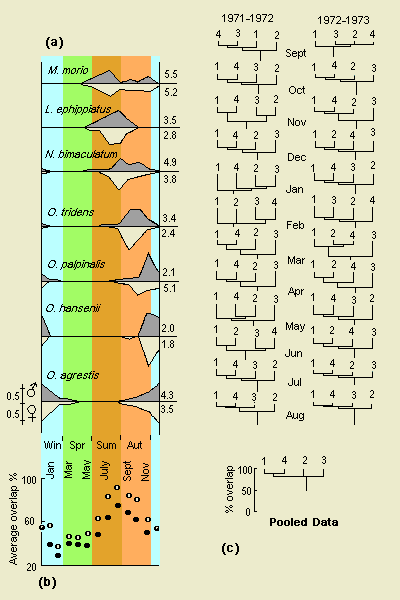
Taxocenes at four sites were most similar when the dominant
species (Nemastoma bimaculatum) was most abundant.
Winter spiders had stenochrone males and eurychrone females. Spiders with both sexes stenochrone occurred in late spring/early summer; summer species were eurychrone.
Harvestmen tended to have females slightly more stenochrone than
males.
This paper is concerned with surface-active species in woodland on Inchcailloch. Some aspects have been described previously (Curtis, 1973, 1975).
Site 1, at the north end of Church Ridge, is fairly dry with an acid brown earth. The main tree cover is Quercus robur. Ground flora is in Group I, dominated by bracken (Pteridium aquilinum) and woodrush (Luzula sylvatica) with some bramble (Rubus fruticosus).
Site 2 is in an alder carr with a poorly drained acid peaty soil. Ground flora fits into Group V, having some R. fruticosus, the mosses Eurhynchium praelongum and Thuidium tamariscinum, honeysuckle (Lonicera periclymenum) and various grasses, with smaller amounts of bracken and woodrush.
Site 3 is near the summit of the North Hill with tree cover mainly P. sylvestris with some birch and rowan (Sorbus aucuparia). Ground flora is of Group X including mainly heather (Calluna vulgaris), blaeberry (Vaccinium myrtillus) and various mosses. The soil is a peaty ranker.
Site 4, in the Central Valley, has tree cover of Q. robur with other species including hazel (Corylus avellana) and birch. R. fruticosus is dominant in the field layer, beneath which is much bare earth. Ground flora is of Group III including Pteridium aquilinum, Endymion non-scriptus (bluebell), grasses such as Holcus mollis, with Eurhynchium praelongum. The soil is an acid brown podzol, much wetter than site 1.
There have been criticisms of pitfall sampling (e.g. Turnbull, 1973: 324). Catches depend on both numerical density and degree of activity, i.e. are a function of activity/density of the animals; this activity/density function probably reflects fairly well the relative ecological importance of the species. Low efficiency of the traps will cause acceptably small disturbance to the sampled population. Possible variation in efficiency is a serious drawback, but if results are treated cautiously they can provide useful indications of community properties. Others have expressed similar opinions, e.g. Williams (1962), Merrett (1976), Uetz & Unzicker (1976). Throughout this paper, phrases such as "numbers of spiders" should be taken as referring to activity/density function, not absolute numerical abundance.
Spiders were identified according to Locket & Millidge (1951, 1953); nomenclature follows Locket, Millidge & Merrett (1974). Opiliones (= Phalangida) are according to Sankey & Savory (1974).
Influent species, contributing 5-50% of site total, occurred as follows at sites 1-4:
| Opiliones:- | ||||
| Oligolophus tridens (C.L.K.) | 297 | 390 | 257 | 110 |
| O. (Odiellus) palpinalis (Herbst) | 50 | 221 | 983 | 3 |
| Lacinius ephippiatus (C.L.K.) | 105 | 138 | 0 | 31 |
| Araneida, Linyphiidae:- | ||||
| Monocephalus fuscipes (Blackw.) | 173 | 108 | 16 | 238 |
Associate species each represent less than 5% of site total. The more common of these are:
| Opiliones:- | ||||
| Mitopus mono (Fab.) | 26 | 52 | 79 | 11 |
| Oligolophus agrestis (Meade) | 41 | 18 | 159 | 26 |
| O. hansenii (Kraepelin) | 5 | 6 | 45 | 3 |
| Araneida, Linyphiidae:- | ||||
| Walkenaera acuminata Blackwall | 62 | 27 | 61 | 21 |
| Gonatium rubellum (Blackwall) | 15 | 4 | 130 | 11 |
| Tapinocyba pallens (O.P. Camb.) | 3 | 8 | 59 | 5 |
| Diplocephalus picinus (Blackwall) | 42 | 3 | 0 | 15 |
| Hilaira excisa (O.P. Camb.) | 0 | 34 | 0 | 0 |
| Centromerus sylvaticus (Blackw.) | 36 | 15 | 22 | 30 |
| C. dilutus (O.P. Camb.) | 15 | 11 | 50 | 7 |
| Macrargus rufus (Wider) | 52 | 3 | 35 | 18 |
| Lepthyphantes alacris (Blackw.) | 10 | 18 | 82 | 19 |
| L. zimmermanni Bertkau | 80 | 73 | 182 | 82 |
| L. tenebricola (Wider) | 41 | 23 | 65 | 44 |
Differences between the four communities are apparent in terms of individual species. A full species list is given by Curtis (1973).
Species diversity may be expressed as the probability of interspecific encounter (Hurlbert, 1971), calculated as PIE = [N/(N - 1)][1 - Σ (ni/N)2], where N is total number captured, ni the number belonging to species i. For sites l-4, values were 0.509, 0.7l2, 0.693 and 0.512. Site 2 had highest PIE value in spite of lower species richness than 1 and 3.
Species richness (Fig. lb) showed a slight increase in spring, contrasting with clear fluctuations in PIE. In spite of fairly constant or declining species richness, PIE rose sharply through autumn owing to decreasing dominance by N. bimaculatum, to maximal levels in winter. Diminished activity led to some low diversity values in early spring, especially at sites 1, 2 and 4. Increasing species richness gave higher diversity in early summer, thereafter declining as N. bimaculatum numbers rose.
Over the two annual cycles, PIE varied with number of individuals trapped (N) according to the following regression equations, all significant:
site 1: PIE=0.76 - 0.001(±0.0002)N; -0.70 (corr. coeff.)
site 2: PIE = 0.78 - 0.001(±0.0003)N; -0.54
site 3; PIE=0.80 - 0.001(±0.0002)N; -0.52
site 4: PIE = 0.72 - 0.002(±0.0005)N; -0.61.
In contrast, species richness appeared independent of N with correlation coefficients -0.16, 0.22, -0.08, -0.04 at the four sites.
Similar conclusions held for logarithmic transformations of N.
Species richness ranged from 4 at site 2 in autumn to 33 at site 3 in early summer. Broadly similar seasonal patterns have been observed in other woodlands, e.g. oak, Wytham Wood (Turnbull, 1960b), beech-maple (Elliott, 1930), and pine (Łuczak, 1959).
Among the common spiders of Inchcailloch, the following sequence appeared. Winter species, e.g. W. acuminata, M. rufus, had stenochrone males and eurychrone females, although the rather less stenochrone males of C. dilutus persisted into spring. In spring, similar patterns were shown by M. fuscipes and T. pallens, but in later spring and early summer both sexes of D. picinus and L. tenebricola were stenochrone. No summer species were stenochrone, L. zimmermanni and L. alacris being eurychrone in both sexes. G. rubellum in autumn showed a trend back towards stenochrone males and had the most eurychrone of females (Is = 11). Progressing into winter, C. sylvaticus showed the typical winter pattern.
The opilionids also showed seasonal succession, but all tended to be stenochrone in activity. Females of L. ephippiatus, N. bimaculatum, 0. tridens and 0. agrestis appeared slightly more stenochrone than males. Only in O. palpinalis were males distinctly more stenochrone than females, while in O. hansenii both sexes have a short period of activity on the ground. The most eurychrone harvestman was M. morio (both sexes).
N. bimaculatum at the four sites showed similar rank order of activity over the months; values of Spearman's rank correlation coefficient comparing sites all exceed 0.87. However, in terms of actual numbers trapped there are significant differences (by χ2) with site 3 showing a greater spread of activity, particularly in early winter.
For the pooled catch, taxocenes of sites 1 and 4 appeared most similar, with 2 and 3 distinct. The relationships vary through the year as shown. Increased similarity from August through October is apparent. During March and April site 3 taxocene is distinct with T. pallens the most numerous species in contrast to M. fuscipes at 1, 2 and 4. During July and August 1972, large numbers of L. ephippiatus at site 2 distinguished that taxocene from the others, but in 1973 this was not so. Increased dissimilarity during winter and spring (December - May) reflects higher species diversity at all sites.
Such correlations may be fortuitous, both sets of data following annual cycles. Much more investigation is required to indicate their significance, if any. Correlations with these factors may be quite complex (Pearson & White, 1964a). Insect populations have been shown to be sensitive to large-scale weather conditions (Wellington, 1957).
Periods of maximal species diversity correspond with low temperature and sparse sunshine. The taxocenes include no species which are stenochrone in summer; summer species tend to be eurychrone. As expected, the taxocenes comprise species adapted for the relatively cool and wet conditions encountered adjacent to the Scottish Highlands.
Spider activity patterns described here fit approximately to the types of phenology observed by Merrett (1969), Pearson & White (1964b), Williams (1962) and A. H. Lavery (pers. comm.) with consistent IS values. Three of the four categories described by Merrett (1969) were seen on Inchcailloch, but no summer stenochrone species, possibly reflecting different climatic conditions at Loch Lomond compared with Dorset.
These patterns of seasonal activity may benefit the community by reducing temporal overlap of otherwise similar species and thus preventing over-exploitation of resources. Climatic factors to which the species are adapted are modified by the annually and spatially varying effect of vegetation, as are actual physical structures afforded to the animals. The moderating effect of vegetation on climate is well known (Geiger, 1965; Cloudsley-Thompson, 1962) and the role played by specific plant structures in affording shelter to invertebrates is considered by Bossenbroek, Kessler, Liem & Vlijm (1977a,b). The physical structure of the vegetation is also important to spiders (e.g. Duffey, 1966). Low shrubs (C. vulgaris, V. myrtillus) at site 3 persist throughout the year, moderating microclimatic changes, and could be partially responsible for the relatively extended period of activity of N. bimaculatum here. At the other sites, most plants die down during late autumn. Some work has examined climatic conditions on Inchcailloch (G. S. Langley, pers. comm.) with fairly predictable results.
The synoptic parameters of species richness and diversity provide some information about taxocene structure, and it is tempting to attempt to relate these to stability, etc. However, for a taxocene which varies so much over the year stability is rather a difficult concept to apply.
PIE may be calculated for a particular species. For N. bimaculatum PIE rises as its abundance declines and falls again as numbers increase. Conversely, the probability of intraspecific encounter rises with increasing activity/density. Cynics may argue that PIE is merely a reflection of the numerical variations and has no biological meaning. However, the probability of intraspecific encounter reaching a maximum may be a partial cause of the subsequent decline in N. bimaculatum as a result of excessive competition, shortage of resources, etc. Clearly, further studies are necessary to clarify this.
|
Figure 1.Inchcailloch spiders and harvestmen: seasonal variations in (a) number of specimens (N) taken per month, (b) species richness (S) and (c) PIE. Symbols relate to sites as indicated by final points in (a). (d) Seasonal activity of more common spiders with males above (shaded) and females below axis; showing proportion of annual total taken in each month. Is values indicated. Back to text |
|
Figure 2.(a) Seasonal abundance of harvestmen with males above and females below axis: showing proportion of annual total taken in each month. IS values indicated. (b) Variations in overlap between the four taxocenes; average of six values between the four sites. (c) Single-linkage cluster analysis dendrograms showing varying relationships of taxocenes at the four sites. Back to text |
 Ariadne Home Page Ariadne Home Page | |