2. Differentiation and 3. Migration of neurons (§2.3)
Unspecialised embryonic neurons (neuroblasts) must differentiate into neurons and also migrate to their 'final' position. These events occur at roughly the same time. Differentiation involves switching on particular genes (induction), which can then be expressed.
How a given cell differentiates depends on its:
(a) lineage - (its 'ancestry'). Time of development is also important; e.g. in rat forebrain, the later developing cells migrate furthest - to the outer cortex.
(b) local environment - the particular signals that the cells receive during development affect their subsequent development; e.g. if early cortical cells are transplanted to a different part of the brain, they develop according to this new position, rather than where they came from originally.
4. Axon growth (§2.4)
Neurons make connections via their axon processes. Axons must first grow and then establish functional connections (synapses) with appropriate targets. Neuroblasts develop several small processes (neurites), one of which becomes the axon (the rest become dendrites). Axonal growth can occur once a neuron reaches its final position or during migration.
Growth cones (the growing tip of an axon) must be stimulated to grow, and also to grow in the right direction towards the 'target'. Several factors are involved:-
Chemotactic factors act locally - helping the axon to 'feel its way';
chemotropic
factors act at a distance ["tropic" = "growing towards"], and the neuron
homes in on this 'beacon'. Distinguish from chemotrophic
(survival) factors ["trophic" = "feeding"] - see below.
Chemotactic factors: Cell adhesion molecules and receptors;
laminin.
Glial cells also play a role (by supplying chemotrophic factors)
Chemotropic factors: Guidepost cells (§ 2.4.4);
"Max factor" (§ 2.4.5)
The timing is important: growth factors are not produced continuously,
and neurons can only respond at certain periods. Some factors appear to
inhibit growth.
Axon guidance is a complex process: axons are affected by their environment
and in turn can affect their environment.
5. Synaptogenesis (Ch 3)
Formation of functional contacts (synapses) with target cells (e.g. other neurons, muscles, etc.) Not all neurons survive. Survival is selective and involves 'matching' neurons with their targets.
Even amongst neurons that reach their targets, there is massive cell
death. Neurons 'die' if they fail to reach a target, or if they fail to
maintain functional contact.(§3.2.1)
To survive, they need:
functional synapses: neural activity is necessary for survival (§
3.1);
chemotrophic (survival) factors (e.g. Nerve Growth Factor) (§
3.2.2);
appropriate targets: neurons that reach the 'wrong' target die (§
3.2.3, 3.2.4). Again, neural activity (action potentials) is important
in the selection process.
6. Refinement of synapses (§ 3.3, 3.4)
Selective pruning of inappropriate connections and strengthening of
functional synapses.
a. Afterent (peripheral) inputs are important (e.g. whisker barrels
§3.3.1: muscle afferents §3.3.2)
b. Selective reduction ('pruning') of axonal branches making
inappropriate projections, e.g. projections from areas of the cerebral
cortex to pyramidal tracts in rats, and refining connections between right
and left visual cortices through the corpus callosum (§3.4.1)
c. Functional tuning to reduce the extent of terminal branches
within target area: e.g. retinal ganglion cell projection to LGN (§
3.4.2).
Neural activity is necessary for this final process, perhaps by selectively
strengthening some synapses at the expense of others (§ 3.4.3) (cf.
memory formation - see later in course)
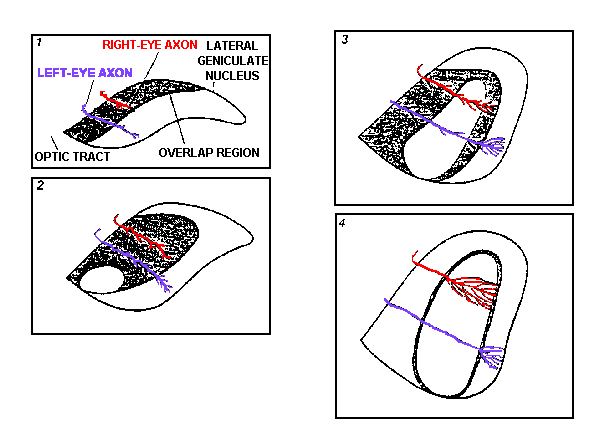
Axonal remodelling in the lateral geniculate nucleus occurs largely before birth. As shown in this figure, at the earliest times in development (1), the axons from the left eye and right eye are simple and tipped with growth cones. The shaded region represents the intermixing of inputs from both eyes. After further development (2), the axons grow many side branches. The axons soon begin to lose some side branches and start to extend elaborate terminal branches (3). Eventually these branches occupy the appropriate territory to form eye-specific layers (4).
See also Fig. 3.9 in Book 4.
There is little evidence that neurons divide in mature mammals; continued
growth and replacement of olfactory neurons is an exception.
Regeneration can occur in the peripheral nervous system (PNS) and central
nervous system (CNS) in fish and amphibia. (§ 3.5.1, 3.5.3) Neuronal
regeneration can occur in mammalian PNS, but not CNS.
The types of glia present are important: Schwann cells (PNS) promote
regeneration; CNS glia (oligodendrocytes) inhibit regrowth.
The intact central part of injured peripheral axons can regenerate
and grow along the glial (Schwann cell) sheaths of the degenerated peripheral
portion. (§3.5.2) Such regrowth does not occur in the mammalian CNS
(§3.5.4). BUT see the recent "Stop Press" from the Course Team.
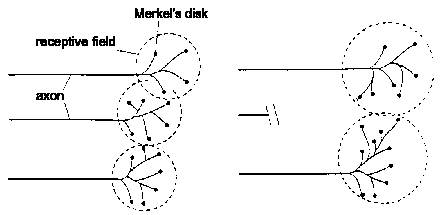
Peripheral axonal damage is accompanied by sprouting of adjacent axon
collaterals, which can take over the denervated target (§3.6.1). In
time, the injured axon may regenerate and reestablish its receptive field.
It has to compete with the adjacent collaterals for target space.
From Fig. 3.14 of Book 4.
Removing peripheral inputs to dorsal horn cells (by cutting dorsal roots
- §3.6.2) results in altered receptive fields of the DH cells - axonal
sprouting and/or unmasking of 'silent' synapses already there.
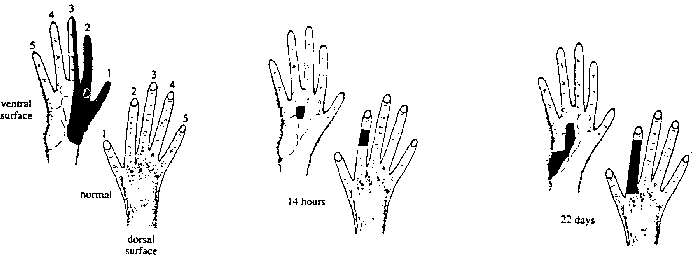
Removing inputs to cortical cells by cutting peripheral nerves (e.g.
median nerve - §3.6.3). Cortical cells no longer respond to stimulation
of original RF. But these neurons soon develop new RFs adjacent to their
original ones - due to axonal sprouting or unmasking of 'silent' synapses.
From Fig. 3.16 of Book 4.
Factors affecting neural development include: genes, external stimuli and hormones. These in turn can influence subsequent behaviour.
Physical differences may arise from:
Differential (non-isometric) growth
'Decision points': external factors having particular
effects at specific times, e.g. nutrition, environmental temperature, humidity,
influence of pheromones.
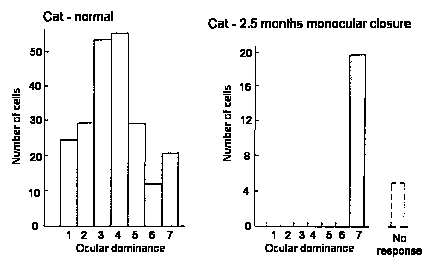
Effects of monocular inputs on ocular dominance columns.
Connections within visual cortex adjust so that normally binocular
cells become monocular. (see below).
Effects of deprivation can be reversed up to a certain developmental stage (e.g. 3/12 in cats).
Similar effects on orientation columns if animals are reared in special
environments devoid of 'vertically' or 'horizontally' oriented 'stimuli'.
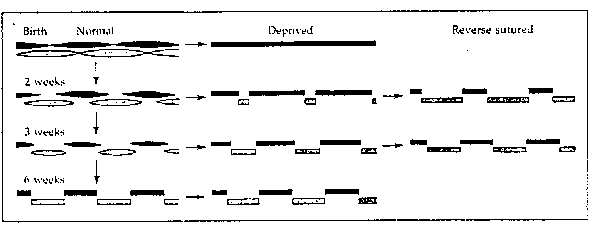
.
SCHEME ILLUSTRATING EFFECTS OF EYE CLOSURE. In the normal cat or monkey
by 6 weeks ocular dominance columns have become well defined in layer IV
of the cortex. Lid closure causes excessive retraction of fibres supplied
by the deprived eye. Those supplied by the open eye retract less than usual
and in the adult supply larger areas of cortex than in normal animals where
the competition is more equal. After reverse suture during the critical
period. the initially deprived eye can recapture the territory it had lost.
(After Hubel and Wiesel, l977.)
In rats and monkeys, testosterone has a maculinising action by preventing
feminisation. (Fetal mammals become female unless exposed to testosterone.)
Without this organising effect, normal male behaviour cannot be activated
at a later stage by testosterone.
Organising effects can cause neural changes. In rat CNS, testosterone
increases the size of the medial preoptic nucleus and amygdala and also
the spinal nucleus bulbocavernosus
Different effects are found in birds. In Japanese quail, testosterone
and oestrogen are both feminising. But in zebra finches, testosterone and
oestrogen are masculising and promote singing by promoting growth of the
higher vocal centre and robustus archistriatalis. (Here, oestrogen is the
organiser, and testosterone the activator.)
These different effects are possible, because testosterone can be converted
to oestrogen inside cells. Unlike peptide hormones, which act on outer
cell membrane receptors, steroid hormones act on receptors inside the cells.
Human development (§4.5) is also subject to similar influences
of genes, external environment and hormones, although the effects can only
be observed, and not tested by experiment (why not?)
or